Functions of Cells and Human Body
Content:
1. Introduction to the gastrointestinal motility
2. Motility of the stomach
3. Motility of the small intestine
4. Motility of the colon
_
Introduction to the gastrointestinal motility
The term motility is defined as involuntary mobility of human tubular organs. To ensure the efficient digestion of food is necessary not only the presence of active enzymes, but also a shift and mixing of chyme during passage through the digestive tube. For this purpose, there are two types of movements in the GIT:
1) Propulsion movements
2) Mixing movements
This division is artificial, because, in fact, both types of movements use the same mechanisms and often one can convert to the other.
Ruchy pędne
Ruchy pędne to ruchy, które zapewniają przemieszczanie się chymu w przewodzie pokarmowym. I to w tempie, które jest proporcjonalne do tempa wchłaniania i trawienia.
Perystaltyka

Podstawowy ruch napędowy nazywany jest perystaltyką. Jej zasada jest prosta: okrężna warstwa mięśniowa w określonym miejscu kurczy się i tworzy pierścień kurczliwy, który jest dalej przesuwany w kierunku brzusznym. W ten sposób powoli popycha on śluzówkę do przodu. Rozszerzenie jelita jest często impulsem do powstania pierścienia kurczliwego – a tym samym do rozpoczęcia perystaltyki. Większa ilość pokarmu powoduje rozciągnięcie i tym samym pobudzenie jelitowego układu nerwowego. Wywołuje to skurcz odcinka mięśnia okrężnego, który znajduje się w odległości kilku centymetrów od miejsca maksymalnego rozciągnięcia przewodu. Perystaltyka jest również wywoływana przez pewne bodźce chemiczne lub silną aktywację przywspółczulną. Występuje ona również automatycznie w określonych odstępach czasu. Obok skurczu dystalnego odcinka przewodu w kierunku doustnym, występuje tzw. relaksacja receptywna w kierunku doustnym od dystalnego odcinka przewodu. Ułatwia to przesuwanie się treści pokarmowej, ponieważ rozluźniona cewka stawia mniejszy opór przesuwającej się treści pokarmowej. Cały ten proces jest kontrolowany przez splot Auerbacha (lub splot jelitowy) i nazywany jest odruchem perystaltycznym.
Ruchy popędowe można eksperymentalnie wywołać również w kierunku ustnym, ale zanikają one po kilku milimetrach. Ruchy perystaltyczne są więc jednokierunkowe, chociaż fizjologiczne podstawy tego zjawiska nie są jeszcze do końca poznane.
Ruchy mieszające
Ruchy mieszające zapewniają stałe mieszanie śliny, tak że cała objętość ważnych odżywczo składników jest wystawiona na działanie enzymów i wchodzi w kontakt z wyściółką jelita w celu wchłonięcia. Ruchy te mają różne formy i różnią się w całym przewodzie pokarmowym.
Segmentacja
Segmentacja to dobrze rozumiany ruch mieszający. Możemy go sobie wyobrazić jako powtarzające się skurcze odległych o kilka centymetrów odcinków mięśnia gładkiego okrężnego. Skurczone regiony różnią się po każdym cyklu segmentacji. W ten sposób następuje segmentacja treści pokarmowej – powstawanie oddzielonych porcji treści pokarmowej, które następnie są ponownie dzielone i jedna połowa jest łączona z porcją poprzednią, a druga z następną. Liczba porcji stopniowo wzrasta, a ich objętość maleje, ponieważ jedna połowa z dwóch zewnętrznych porcji zawsze nie ma gdzie się przyczepić i tworzy nową porcję.
_
Ruchliwość żołądka
Żołądek dzięki układowi swoich mięśni jest dobrze wyposażony do pełnienia trzech funkcji:
1) Mieszanie pokarmu
2) Przechowywanie dużych ilości pokarmu
3) Opróżnianie do dwunastnicy
Mieszanie pokarmu
Do czasu obecności śluzu w żołądku, w jego górnej 1/3 powstają słabe fale skurczowe (zwane falami mieszania). Pojawiają się one regularnie co 20 sekund i opierają się na automatyzmie pracy mięśni gładkich. Gdy fala mieszania rozciąga się od trzonu żołądka do antrum, staje się silniejsza i silnie popycha śluzówkę do odźwiernika. Jednakże, jak zwieracz odźwiernika jest zamknięty, napotyka chyme do ślepych zakończeń i tylko mieszać, odpowiednio, pod wysokim ciśnieniem ucieka z powrotem przeciwko ruchowi pierścienia kurczliwego. Zjawisko to nazywane jest retropulsją.
Dobrze jest zauważyć, że zwieracz odźwiernika nigdy nie jest całkowicie zamknięty. Chym, który przez niego uchodzi, ma objętość zaledwie kilku mililitrów, ale pełni ważną funkcję, gdyż dwunastnica może badać jego skład i według tego regulowana jest motoryka żołądka. To zjawisko, które jest dodatkiem do mieszania retropulsji, nazywamy pompą odźwiernikową.
Opróżnianie żołądka
Opróżnianie żołądka ma taki sam mechanizm jak retropulsja. Jednocześnie jednak towarzyszy mu zmniejszenie oporu przeciwko przechodzeniu treści pokarmowej przez odźwiernik – dochodzi do rozluźnienia zwieracza odźwiernika. Dzięki temu do dwunastnicy może przedostać się określona objętość pokarmu – w zależności od oporu odźwiernika w danym momencie. Ogólnie mówiąc: im wyższe jest napięcie zwieracza, tym większy jest opór przeciwko przechodzeniu chymu przez odźwiernik i tym mniejsza objętość chymu dostaje się do dwunastnicy.
Zwieracz odźwiernika to tak naprawdę tylko grubsza warstwa zwykłego mięśnia okrężnego. Ma on około podwójną grubość w odźwierniku w porównaniu z resztą żołądka. Normalnie jego ton jest ustawiony tak, że opór jest wystarczająco mały dla przejścia płynów, ale zbyt duży dla przejścia stałego chyme. Musi on przejść przez kilka mieszania retropulsji i być wystarczająco zmieszany z sokami żołądkowymi, aby kontynuować do dwunastnicy.
Opróżnianie żołądka jest kontrolowane przez kilka czynników, które można podzielić na dwie grupy:
1) Czynniki żołądkowe
2) Czynniki dwunastnicze
Faktory żołądkowe
Faktory żołądkowe generalnie potęgują opróżnianie żołądka poprzez zwiększenie częstotliwości fal mieszania lub poprzez zmniejszenie napięcia odźwiernika. Uaktywniają się one, gdy w żołądku znajduje się większa ilość pokarmu (a zwłaszcza pokarmu bogatego w białka, np. mięsa). Kontakt peptydów z błoną śluzową antrum powoduje wydzielanie hormonu żołądkowo-jelitowego – gastryny.
Gastrin has the following effects:
1) Increases the production of gastric juice that has low pH
2) Increases the frequency of spontaneous motor activity of the stomach (mixing waves)
3) Decreases the pyloric sphincter tone
Note that if there is a sudden increase of the frequency of mixing waves and a decrease of pyloric sphincter tone, increased efficiency of the pyloric pump occurs. This is the main mechanism of increased gastric emptying.
Duodenal factors
These are mostly inhibitory signals that block gastric emptying. There are two main groups:
1) Nerve feedback to enterogastric system
2) Feedback control through the gastrointestinal hormones
Nerve feedback to enterogastric system
If large volume of chyme passes through the pyloric sphincter into the duodenum, there is a distension of its wall leading to a reflex that slows down or completely stops gastric emptying. This signal is mediated:
1) Directly by the enterogastric system
2) Through the paravertebral sympathetic ganglion
3) Through the vagus nerve to the brainstem and back
All of them are called pyloric reflexes. Their effect is dual:
1) Decreased frequency of mixing waves
2) Increased tone of the pyloric sphincter
This slows down the mechanism of the pyloric pump.
In addition to the volume of the chyme, the pyloric reflexes are activated by the low pH (3.5-4), wysokie stężenie peptydów w ślinie oraz jej hipertoniczność lub hipotoniczność.
Kontrola zwrotna poprzez hormony przewodu pokarmowego
Komórki nabłonkowe dwunastnicy wykazują aktywność sensoryczną dla niektórych rodzajów składników pokarmowych. Aktywują one odruchy odźwiernikowe przy zmianach pH, stężenia peptydów lub toniczności. W obecności lipidów przyspieszają wydzielanie tych mediatorów (nie tylko w obecności lipidów, ale na lipidy są najbardziej wrażliwe):
CCK – cholecystokinina
Hormon ten ma trzy ważne działania. Zmniejsza aktywność pompy odźwiernikowej, blokuje wpływ gastryny na motorykę żołądka i zwiększa wydzielanie żółci do żółci, ponieważ aktywuje skurcze pęcherzyka żółciowego.
Sekretyna
Sekretyna jest produkowana przez komórki dwunastnicy w odpowiedzi na niskie pH treści pokarmowej. It inhibits gastric emptying.
GIP – gastric inhibitory peptide
GIP is produced as a response to the high lipid content in the chyme. Although it has an inhibitory effects on the gastric motility and especially on the the pyloric pump it is the weakest one of all three hormones.
_
Motility of the small intestine
Contractions of the muscle layers of the small intestine can be divided into two groups:
1) Segmentation contractions
2) Propulsion contractions
Segmentation contractions
Process of the segmentation has already been discussed above. We only briefly discuss its causes here. Segmentation is a manifestation of electrical slow-waves, which represent action potentials generated by the automaticity of smooth muscle. Maximal frequency of these slow waves is 12/min. Dlatego segmentacja może występować nawet do 12 razy na minutę, ale tylko w bardzo rzadkich przypadkach. Normalna częstotliwość ruchów segmentacyjnych wynosi około 3 na minutę.
Skurcze pędne
Mechanizm ich powstawania został już opisany w części wstępnej. Pierścień kurczliwy w jelicie cienkim ma prędkość około 0,5-2 cm/min. Szybciej w segmentach proksymalnych, w segmentach dystalnych zwalnia. Jeden pierścień kurczliwy pokonuje maksymalną odległość 10 cm, po czym wychodzi, a błona śluzowa czeka na nowy. Dlatego ogólna prędkość pasażu śluzu wynosi 1 cm/min.
Kontrola motoryki jelita cienkiego
Po każdym posiłku w jelicie cienkim dochodzi do nasilenia ruchów propulsyjnych. Wynika to zarówno z obecności chymu w jelicie cienkim, jak i z odruchu żołądkowo-jelitowego. Odruch ten jest odpowiedzią na rozciąganie ściany żołądka i powoduje wzrost motoryki jelit. Komponenty tego odruchu leżą całkowicie w splocie mięśniowym. Ponadto działają hormony – CCK, gastryna, insulina, motylina. Są one wydzielane poposiłkowo i zwiększają częstotliwość ruchów propulsyjnych i mieszających. Odwrotnie, sekretyna i glukagon hamują motorykę jelita cienkiego.
Zastawka jelitowo-jelitowa
Funkcją tej zastawki jest zapobieganie refluksowi treści pokarmowej z jelita grubego do jelita cienkiego. W rzeczywistości nie jest to zastawka, ale otwór końcowego odcinka jelita krętego, który wystaje do kątnicy. Jednak ze względu na ilość mięśni działa jak zastawka. Jego działanie zależy od jego oporu. Przechodzi przez nią około 1500-2000 ml/dobę śluzu.
_
Motoryka jelita grubego
Jelito grube ma dwie główne funkcje: wchłanianie elektrolitów i wody oraz magazynowanie odpadów stałych przed ich eliminacją z organizmu. Obie te funkcje nie wymagają rozległej aktywności ruchowej. Dlatego warstwa mięśni podłużnych jest w okrężnicy zredukowana do taenii. Stanowią one trzy pasma mięśniowe, które rozciągają się na całej długości okrężnicy. Jego motoryka jest więc wolniejsza niż jelita cienkiego.
Haustracja
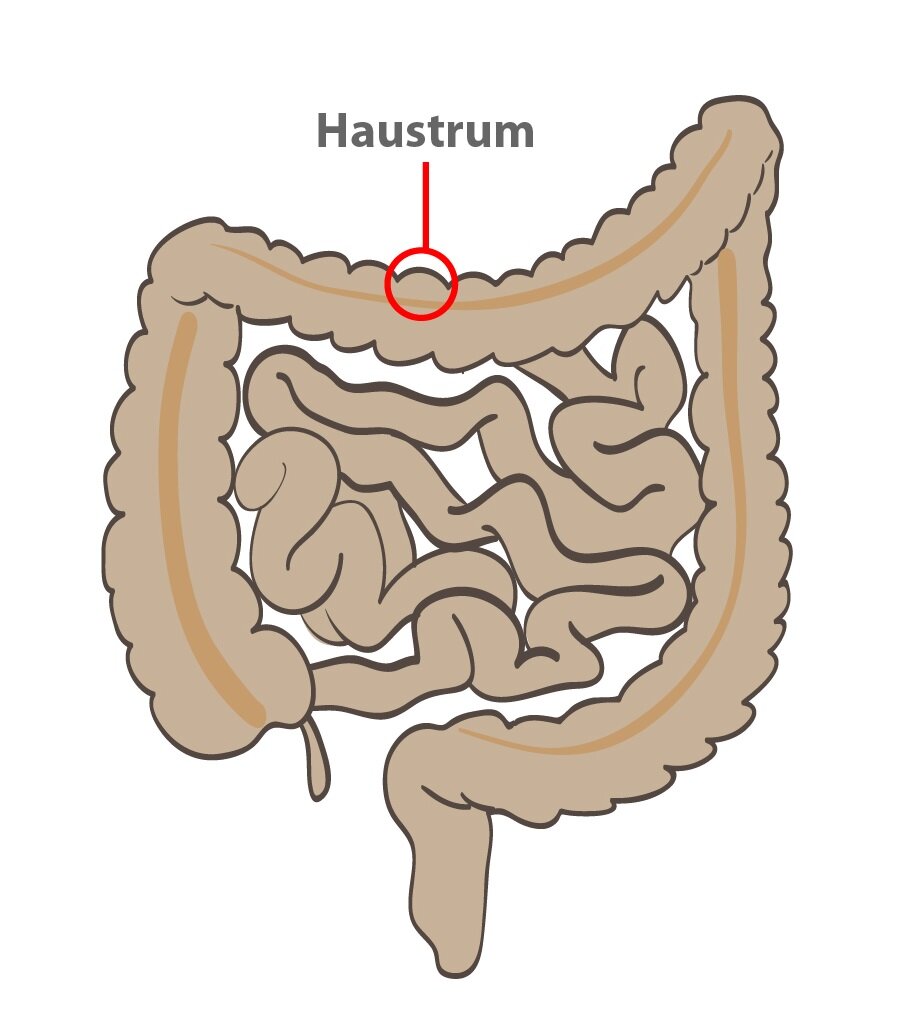 Jest to ruch mieszający jelita grubego (zmodyfikowany ruch segmentacyjny). Najpierw dochodzi do skurczu mięśni okrężnicy. Następnie kurczą się taeniae tworząc haustrum. Haustra tworzą charakterystyczny wygląd okrężnicy, która składa się z szeregu stosunkowo dużych wybrzuszeń. Podczas skurczów taenii wewnątrz haustra występuje zwiększone ciśnienie. Po około 30 sekundach ciśnienie osiąga wartość maksymalną, a w ciągu następnych 60 sekund haustrum znika. Tworzenie nowego haustrum rozpoczyna się po kilkunastu minutach w nowym miejscu (ruch segmentacyjny), a cały proces powtarza się.
Jest to ruch mieszający jelita grubego (zmodyfikowany ruch segmentacyjny). Najpierw dochodzi do skurczu mięśni okrężnicy. Następnie kurczą się taeniae tworząc haustrum. Haustra tworzą charakterystyczny wygląd okrężnicy, która składa się z szeregu stosunkowo dużych wybrzuszeń. Podczas skurczów taenii wewnątrz haustra występuje zwiększone ciśnienie. Po około 30 sekundach ciśnienie osiąga wartość maksymalną, a w ciągu następnych 60 sekund haustrum znika. Tworzenie nowego haustrum rozpoczyna się po kilkunastu minutach w nowym miejscu (ruch segmentacyjny), a cały proces powtarza się.
Ruchy napędowe
Ruchy napędowe są w większości zdeterminowane przez haustrację, która stopniowo i powoli postępuje od jelita ślepego do esicy. Przejście strawionego materiału przez okrężnicę w wyniku haustracji trwa około 12 godzin, a płynna treść pokarmowa staje się stałą masą kałową.
Jednakże istnieje szybszy ruch napędowy okrężnicy, który występuje trzy razy dziennie przez około godzinę po posiłku i trwa tylko 15 minut. Przypomina on perystaltykę. Istnieje pierścień skurczowy, który stopniowo przesuwa się w kierunku brzusznym. Powstaje on w okrężnicy poprzecznej i normalna czynność przełyku zanika na około 15 minut.
Modulacja motoryki jelita grubego
Wszystkie opisane ruchy jelita grubego zwiększają swoją intensywność i częstotliwość po wyzwoleniu dwóch odruchów:
1) Gastrocolic reflex
2) Duodenocolic reflex
Gastrocolic reflex
Gastrocolic reflex is triggered by a high tension in the stomach wall. Myenteric plexus transports this signal through to the colon that increases the frequency of haustra formation.
Duodenocolic reflex
Duodenocolic reflex is triggered by a high tension in the duodenal wall. Signal spreads through the myenteric plexus to the colon and increases the frequency of action potentials in the smooth muscle cells. That increases speed of the propulsion movements.
Subchapter Author: Patrik Maďa
![]()